

İŞİTME FİZYOLOJİSİ VE FİZYOPATOLOJİSİ
İşitme fizyolojisini anlatmadan önce ses hakkında genel bilgi vermek konunun anlaşılması yönünden verimli olacaktır.
Ses: Dış ortamdaki moleküllerin longitudinal titreşimlerinin kulak zarına çarpması ile meydana gelen bir duyudur. Sesin meydana gelebilmesi için bir cismin titreşmesi, titreşen bu çişimin de hava veya, su gibi bir ortamda bulunması şarttır.
Frekans: Saniyedeki titreşim sayısı veya bir zaman birimindeki dalga sayısıdır. Bir zaman dilimindeki titreşim sayısı ne kadar çoksa ses o kadar tizdir, frekans sayısı azaldıkça ses o kadar pesleşir. Normal işiten insan kulağı 16 Hz-20000 Hz arasındaki sesleri algılamaktadır.
Şiddet: Ses dalgalarının amplitudine bağlıdır. Bir sesin şiddeti kulak zarındaki değişikliklerle ölçülebilirse de relatif bir ölçü kullanmak daha iyidir. Bu nedenle işitmederecesi Desibel (dB) ile ölçülmektedir.
Tını:Sesin kapsadığı katmer titreşimlere bağlıdır. Müzikal seslerin çoğu sesin yüksekliğini tayin eden primer bir frekans ile birçok harmonik titreşimlerden meydana gelir ki bu sonuçlar sese karakterini yani tınısını verirler.
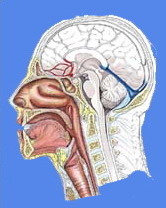
DIŞ KULAK FİZYOLOJİSİ
Dış kulak işitme olayında pasif bir rol oynar. Baş ve vücut ile birlikte boşlukta sesin yayılmasını etkiler ve sonuç olarak akustik sinyallerin timpanik zara ulaşmasına etki eder. Baş, ses sahasında bir engel olarak hareket eder. Baş, dalgaları yüzeyinden yansıtır, aynı zamanda ses dalgalarının kırınımına neden olur.
Baffle etki: Baş gibi bir yüzeye çarpan ses dalgalarının yansıtılacağını ifade eder. Yüzeye çarpan ve yansıyan dalgalar başın yan tarafında daha büyük bir ses basıncı oluşturacak şekilde birleşirler.
Shadovv etki: Başın boyutu ile karşılaştırıldığında relative olarak kısa dalga boyuna sahip ses dalgaları bir ölçüde bloke edilir ve karşı tarafta daha düşük ses basıncı ile sonuçlanır. Shadovv etki sesin frekans veya dalga boyuna bağlıdır. Yüksek frekanstaki ses başın daha çok shadovv etki yapması ile sonuçlanır ve sonuç olarak başın her iki tarafındaki ses lokalizasyonu için daha büyük interaural yoğunluk farkı oluşur.
Kulak kanalı girişi ve kulak kanalı, akustik rezonatörler olarak hareket eder. Böylece timpanik zarda ses basıncı etkisi yaparlar. Kulak kanalı bir ucu açık basit bir tüp gibi hareket eder. Böyle bir tüpün boyu, büyük ölçüde rezonant karakteristiklerini belirler. Tipik olarak rezonant pik noktası 3.5 kHz kadardır.
Baş, vücut ve external kulağın birkaç özelliğinin sesin lokalizasyonunu sağlamada önemli olduğu düşünülür. Yüksek frekanslı stimulus için baffle ve shadovv etkisinden kaynaklanan interaural yoğunluk farkı boşluktaki ses kaynağının pozisyonuna bağlıdır. Başın yanındaki kaynaklar stimulus yoğunluğunda büyük farklılıklara yol açabilir. Aynı zamanda iki kulak arasındaki uzaklık bir kulaktan diğerine ses ulaşımı arasında zaman farklılığına yol açar. Bu interaural zaman farkı aynı zamanda boşluktaki ses lokalizasyonuna bağlıdır. Baş etrafındaki ses kırınımı, lokalizasyona katkıda bulunabilir. Bir cisim etrafındaki farklı pozisyondaki ses kaynakları koronal plan civarında baş ve kulak kepçesinin asimetrisinden dolayı farklı kırınım paternlerine neden olur. Serbest alandan kulak zarına ses basıncı transformasyonu, ses istikamet açıları değiştiğinden farklı frekans bağımlılığı gösterir. Böylece baş ve dış kulak, ses kaynağının farklı pozisyonları için farklı bir tarzda timpaniuma ulaşan geniş bir inyal bandının spektrumuna etki eder.
ORTA KULAK FİZYOLOJİSİ
Orta kulak, akustik enerjiyi hava ile dolu external kulak kanalında, sıvı ile dolu kohleaya iletir. Normal bir orta kulak şu yapılardan oluşur: Kendisine gelen ses titreşimlerini alan bir kulak zarı, bu titreşimleri iç kulağa ileten solid bir sistem yani kemikçik zinciri, normal çalışan pencereler ve zarın her iki tarafındaki hava basıncını dengeleyen tuba östaki orta kulak, hava ve sıvı ortamlar arasındaki impedans ayarını yapar.
İmpedanstan biraz bahsedecek olursak sesin bir ortamda yayılması ortamın elastisitesine vöe yoğunluğuna bağlıdır, rezistans ile impedans aynı anlamdadır. Akustik rezistans, ortamın ses dalgalarının yayılımına gösterdiği dirençtir. Suyun akustik rezistansı çok büyüktür.(160 000) Havanın akustik rezistanıs ise çok küçüktür(42). Hava ve kohlear sıvı arasında oldukça büyük bir impedans farkı mevcuttur. Havadan gelen sesin büyük bir kısmı orta kulak olmasaydı geri yansırdı. Bu durum impedans farkından kaynaklanır. Eğer impedanslar yakın hale getirilirse ses enerjisi bir ortamdan diğerine olduğu gibi geçirilir. Enerji kaybı olmaz. Buna impedans ayarlaması denir. Orta kulak havanın düşük impedansını, iç kulak sıvısının yüksek impedansma ayarlayarak bir transformer görevi görür. Eğer ses dalgaları direkt olarak kohlear sıvıya ulaşsalardı 30 dB'lik bir kayba uğrarlardı. Orta kulak bir çeşit amplifikatördür. Sesi iç kulağa geçerken 30 dB daha kuvvetlendirir.
İmpedans uyumu üç yolla başarılır 1.)İlk ve en önemli faktör timpanik membranın efektif titreşim alanının, stapes tabanının efektif titreşim alanından 17-20 kez daha büyük olmasıdır. 2.) Kemikçik zincirinin kaldıraç hareketini içerir. İnkusun uzun kolu, malleusun boynu ve manibriumun uzunluğundan 1.3 defa daha kısadır. 3.) Timpanik membranın şeklidir. Kuşlar ve diğer omurgalılarla karşılaştırıldığında eğimli bir koni şeklinde olan memeli zarı, daha seçici ve daha geniş frekansları algılar niteliktedir. Bu üç faktörün kombine etkisi sonucu 30 dB'e yakın basınç kazancıdır.
İç kulağa güç transferine ek olarak timpanik membran, orta kulağı kulak kanalı yabancı materyalinden korur ve nazofarenksten östaki tüpü vasıtası ile yabancı materyalin içeriye üflenmesini önleyen hava yastığını sürdürür.
Baş, dış kulak, orta kulağın akustik özellikleri ve aynı zamanda kohleamn input impedansmın kombine etkileri işitme fonksiyonu üzerinde çok önemli bir role sahiptir.Bu faktörler işitme eğrisinin şeklini ve bu nedenle de insanın işitme frekansının range'nı belirler. Diğer bir ifade ile insanların 20 kHz veya daha yukarı sesleri takip edememe ve tanıyamamalarının bir nedeni, böyle yüksek frekanslı seslerin etkin bir şekilde orta kulak ve kohleaya iletilemeyişidir.
Timpanik membnanın titreşimi çeşitlilik gösterir. Birçok araştırmacı timpanik membranın dinamiğini incelemiştir. Kulak zarının titreşme amplitüdleri çok düşük olup, çıplak gözle ve ameliyat mikroskopu ile görülemez. 2000 Hz'lik bir sesle kulak zarının titreşim amplitüdünün 0.0045 u olduğu saptanmıştır ki bu büyüklük hidrojen molekülünün yapısının yarısına eşittir.
Ostaki tüpü fonksiyonlarına kısaca değinmek gerekirse
1*. Orta kulağı nazofaringeal sekresyonlardan korumak.
2*. Orta kulaktaki sekresyonları nazofarenkse drene etmek (Bu fonksiyon tubadaki mukosiliar aktivite ile sağlanır).
3*. Ventilasyonu sağlayarak orta kulaktaki hava basıncını dışarıdaki atmosferik basınca eşitlemek. İdeal tüp fonksiyonunun varlığında istirahat halinde tuba kapalıdır. Yutma esnasında ise M. tensor veli palatini'nin kasılması ile açılır.
Kemikçik zinciri ses enerjisini mekanik enerjiye çeviren bir transformerdir. Titreşimleri zardan kohleaya aktarmaktadır. Kemikçik zincirinin önemli bir özelliği, ses titreşimlerinin stapes tabanına ve yuvarlak pencereye aynı anda gitmesini önlemesidir. Kulak zarının titreşimleri , hem kemikçikleri hem de orta kulak boşluğundaki havayı titreştirir. Yuvarlak pencereye ses titreşimleri orta kulaktaki hava ile, oval pencereye ise kemikçik zinciri ile gelir. Her iki ses titreşimi arasında iletim hızı yönünden bir faz farkı ortaya çıkar. Bu olaya dezafaj denir. Waver ve Lavvrance'nin bu konuda yaptığı çalışmalar göstermiştir ki hem oval hem de yuvarlak pencereye ses dalgaları farklı fazda gönderilirlerse elde edilen kohlear potansiyel en yüksek, aynı fazda gönderilirlerse en düşük seviyede bulunmaktadır. Bunun nedeni, aynı fazda gönderilen ses dalgalarının perilenfe aynı yönde hareket etkisi yapmaları ve bu yüzdende her iki dalganın birbirini yok etmesi; bunu karşın farklı fazda gönderilen dalgaların ise birbirine aksi yönde hareket etkisi yaparak amplitüdü yükseltmeleridir. Normal bireylerde, yani kulak zarı ve kemikçikler sistemi sağlam kişilerde bunun önemi azdır. Çünkü ses dalgaları kemikçikler yolu ile kuvvetlenerek iç kulağa geçerler. Oysa orta kulak havasının titreşimleri ile yuvarlak pencereye gelen ses dalgaları bu olanaktan yoksundur. Orta kulağın amplifikatör etkisinden yararlanamaz ve 30 dB'lik bir kayıp ile iç kulağa geçebilir. Pencerelerin asıl önemi kulak zarının perfore ve kemikçikler sisteminin eksik olduğu zamanda ortaya çıkar. Eğer ses dalgaları, kulak zarı perforasyonu ve kemikçikler sisteminin eksikliği nedeni ile hem yuvarlak hem de oval pencereye hava yolu ile geçerse yani her iki pencereye aynı fazda ulaşırsa bu zaman perilenfe aynı yönde hareket etkisi yapacaklar ve sıvıların hareketi birbirini yok edecek ve işitme düşecektir. Eğer pencerelerden biri korunacak olursa ses dalgalarının aynı fazda her iki pencereye ulaşma olanağı kaldırılacak ve perilenfe yerdeğiştirme olanağı sağlanacaktır.
Kulak zarı perforasyonu ses iletimini engeller, bu engellenme şu nedenlerle olur: Perforasyon yüzünden titreşen kulak zarı yüzölçümü küçülür. Orta kulağın amplifikatör etkisini yapan kulak zarı ve oval pencere yüzölçümleri arasındaki oran küçülür. Perforasyon kenarlarına çarpan ses dalgaları iki ayrı dalga oluşturur: Kulak zarı dış yüzünde ve kulak zarı iç yüzünde. Bu iki dalga zıt yönlü oldukları için birbirlerinin etkisini ortadan kaldırır. Eğer perforasyon büyükse hem oval hem de yuvarlak pencere aynı anda ses dalgalarını alır, dezafaj olayı ortadan kalkar.
Orta kulakta iki düz adele Tensor timpani ve Stapedius adelesi yer almaktadır. Tensor timpani kası malleusa tutunur ve trigemindl sinir tarafından innerve edilir. Stapedius adelesi ise stapese tutunur ve fasial sinirin stapedial dalı tarafından innerve edilir. Stapedius ve tensor timpani adelelerinin her ikisi de, sadece vücuttaki en küçük düz adele oldukları için değil, aynı zamanda oldukça yüksek innervasyon oranına (adele lifine isabet eden sinir lifi sayısı olarak) sahip oldukları için de önemlidirler. Bu adelelerin kontraksiyonunun orta kulaktaki ses transformasyonuna etkileri konusunda şüphe yoktur. Fakat halen bu etkinin detayları ve orta kulak kaslarının ne ölçüde etkili olduğu konusu tam olarak anlaşılmış değildir. Aslında orta kulak adelelerinin kendiliğinden yaptığı daha birçok fonksiyonunda olduğu söylenmektedir.
Orta kulak adelelerinin bir fonksiyonuda kohleayı yüksek seslerden korumaktır. Ses, 80 dB'den fazla olduğu zaman Stapedius adelesinin otomatik olarak reflex kontraksiyonu vardır. Bu kontraksiyon ossiküler zincirin ve timpanik membranın gerginliğini arttırır. Böylece sesi 2 kHz'in altına indirir. Gürültü etkisi ile işitme kaybı konusundaki laboratuar ve saha araştırmaları özellikle 90 dB'i geçen, düşük frekanslı seslerde (2 kHz'den az) stapedial refleksin kohleayı koruduğunu göstermektedir. Akustik refleksin latensi 10 msec'ten daha büyük olduğu için kohlea, beklenmeyen kısa süreli impulsif seslerden korunmayabilir.
Orta kulak kaslarının diğer fonksiyonları ise ossiküler zincire gerginlik ve rijidite sağlamak, ossiküler zincirin beslenmesine katkıda bulunmak, çiğneme ve vokalizasyonun sebep olduğu fizyolojik gürültünün azaltılması, yüksek seviyedeki düşük frekanslı seslerin hafifletilmesidir.
İÇ KULAK FİZYOLOJİSİ
İç kulak, temporal kemik içerisine yerleşmiş olup,zar ve kemik yapılardan oluşur. İnsan kohleası halka şeklinde ve yaklaşık 35 mm. boyunda scala vestibuli, scala media ve scala timpani bölümlerine ayrılmış kemikli bir tüptür. Vestibuli ve timpani bölümleri, ekstrasellüler sıvı benzeri ve içinde 4 mEq/lt K iyonu ile 139 mEq/lt Na iyonu bulunan perilenf denen bir materyal içermektedir. Scala media, ReissnerS membranı, Basilar membran, lamina spiralis ossea ve lateral duvar ile çevrilmiş olup, endolenf içermektedir ki bu materyal de intrasellüler sıvı benzeridir.İçerisinde 144 mEq/lt K iyonu ile 13 mEq/lt Na iyonu konsantrasyonuna sahiptir. Scala media yaklaşık 80 mV pozitif DC'ye (Doğru akım) sahiptir ki bu akım tabandan apekse doğru hafifçe azalmaktadır. Bu endokohlear potansiyel, oldukça vaskülarize olan kohleanın lateral duvarının stria vaskülarisi tarafından üretilmektedir. Stria vaskülarisin özelleşmiş birçok hücresi Na/K ATP ase pompalar.
Oval penceredeki stapes tabanının piston gibi hareketi ile kohleaya akustik enerjiyi girmektedir ve doğruca scala vestibulinin perilenfi ile birleşir. Scala vestibulinin perilenfi kohleanm apeksinde Helicotrema denilen küçük bir açıklık yolu ile scala timpaninin perilenfi ile iletişim sağlar. Corti organı, basiler membran ve lamina spiralis ossea üzerinde uzanmaktadır. Basillar membran, tabanda yaklaşık 0.12 mm, apekste genişleyerek 0.5 mm'ye kadar artmaktadır. Corti organının başlıca komponentleri, dış ve iç hair cell'ler, destek hücreleri (Deiter's,Hensen's, Claudıus's destek hücreleri), tektorial membran ve retiküler lamina-Cutikular plate kompleksidir. Destek hücreleri, corti organına yapısal ve metabolik destek sağlar. Ayrıca Deiter's hücrelerinin phalangeal proçesi retiküler laminanın sıkı hücre bağlantılarını oluşturur.
Corti organının dış ve iç hair cell'leri mekanik (akustik) enerjinin elektrik (Nöral) enerjiye transdüksiyonunda önemli rol oynarlar. Dış hair cell'ler, İç hair celilerden oldukça farklıdır, İç ve dış hair cell'ler arasındaki morfolojik farklara ilaveten nöral innervasyon da önemli derecede farklıdır. Dendrit, lamina spiralis osseus'a projekte olurken, aksonları beyin sapının kohlea nükleusuna gönderir.
Kohleanın hareketini stimüle eden 50 000 nörondan %_90-95 kadarı doğrudan iç hair cell'ler ile sinaps yapar ve bunlara tip l nöronlar denir. Herbir iç hair celi 15-20 tane tip ! nöron tarafından innerve edilir. Buna karşın 50 000 nöronun % 5-10 kadarı dış hair celileri innerve eder. Bunlara tip II nöronlar denir. Herbir tip II nöron 10 kadar dış hair cell'i stimüle etmek için dallara ayrılır. Kohleanm afferent innervasyon paternine ilaveten ipsi ve kontralateral superior olivary komplexten kaynaklanan 1800 kadar efferent lif kohleaya yönelmektedir.
Akustik enerji nedeni ile stapesin hareketine cevap olarak basiller membranın hareketi ile transdüksiyon işlemi başlamaktadır. Basiller membranın yer değiştirme paterni gezen bir dalgadır ve dalga hareketi daima tabandan apexe doğru ilerler. Basiller membran hareketinin piki yada maksimum amplitüdü stimulus frekansının bir fonksiyonu olarak değişim gösterir. Örneğin 10 kHz gibi yüksek frekanslı sesler tarafından üretilen dalga hareketi kohleanın tabanına yakın maksimum harekete sahip olduğu halde, 125 Hz gibi düşük frekanslı dalgalar apikal bölgede maksimuma sahiptirler. Yüksek frekanslı ses dalgaları kohleanın apikal bölgesine ulaşmazlar. Fakat düşük frekanslı ses dalgaları basiller membranın tüm uzunluğu boyunca gezebilirler. Son zamanlarda hassas kayıt ve inceleme metodları kullanılarak elde edilen veriler göstermiştirki, kulağın belirgin frekans seçici yeteneklerinin çoğu kohleanın mekaniksel özellikleri ile açıklanabilir. Mekanik dalga hareketinin hassas ayar piki için oluşan mekanizma, kohleanın lokalizasyonuna göre en iyi frekansa yakın frekanslarde, basillar membranın hareketini arttıran dış hair celilerin faaliyetlerine de kapsar. Bu arttırma işlemine kohlea yükselticisi de denir ki hem kulağın ince frekans seçicilik yeteneğine, hem de kulağın hassasiyetine ve çok yavaş sesleride izleme yeteneğine katkıda bulunur. Kohlea yükselticisine katkıda bulunan faktörler, dış hair cell'lerin motolitesi, stereosilyanın mekanik özellikleri ve tektorial membrandır. Stereosilya-hair cell kompleksi transdüksiyon işleminde önemlidir. İç hair cell'lerin stereosilyaları muhtemelen tektorial membran ile temas etmezler. Fakat dış hair celiler doğrudan temas halindedirler. Dalga hareketi tarafından stereosilyanın defleksiyonu duyu hücrelerine K iyon akımını sağlayan stereosiiyalarm uçlarındaki nonspesifik iyon kanallarını açar ve kapatırlar. K iyonlarının duyu hücrelerine akması stereosilyanın iyon kanallarının açılması ve kapanması ile ayarlanır. K akımı, negatif intrasellüler hair celi potansiyellerine +80 mV endokohlear potansiyel ilavesi ile başlar. (-) intrasellüler potansiyeller, iç hair celi için -45 mV ve dış hair celi için -70 mV kadardır. Sonuçta oluşan intrasellüler depolarizasyon kalsiyum içeren enzim salgılanmasına sebep olur ki sonuçta kimyasal transmitterler salıverilmesine ve nuhayet afferent sinir liflerinin aktivasyonuna sebap olur. Afferent sinapstaki spesifik nörotransmitterler bilinmemektedir.
Gross kohlear potansiyeller: Kohleada dört gross (ekstrasellüler) potansiyel kaydedilebilir: Endolenfatik (endokohlear) potansiyel(EP), kohlear mikrofonik (CM), sumasyon potansiyel (SM), tüm sinir aksiyon potansiyeli (ÇAP)
Diğer kohlea potensiyellerinden farklı olarak endolenfatik potansiyel (EP) akustik stimulasyona cevap olarak ortaya çıkmaz.EP daha ziyade scala mediada kaydedilmiş 80-100 mv DC potansiyelidir.EP kohleanın lateral duvarının stria vaskülarisinden çıkar. Stria vaskülaris kohleanın enerji kaynağı olarak kabul edilir ve transdüksiyon işlemi için çok önemlidir. Enerji kaynağının tabiatı stria vaskülarisin yoğun vaskülaritesi ile ilgilidir ve aynı zamanda Na-K ATP’ase ile ilişkilidir. Na-K ATP’ase; stria vaskülarisin marjinal hücreleri, dış sulkus hücreleri, Reissner^s membranına bitişik fibrositler ve spiral ligamentler dahil kohlea hücreleninin çeşitli tiplerinde lokalizedir.
Endolenf ve EP.'nin üretimini kapsayan mekanizmaların düzensiz çalışması bazen metabolik presbycusis denilen işitme kaybı yapabilir.
Kohlea mikrofonik (CM) bir alternatif voltajdır. Genellikle kohlea içinde veya yuvarlak pencerenin yakımda kaydedilir. CM özellikle dış hair celilere K akımını temsil eder ve böylece basiller membranınhareketi ile dış hair celilerin elektriksel düzeni değiştirilir. Stereosilya modiolustan dışa doğru büküldüğünde hair celi direnci azalır ve elektrik akımında artış ve EP'de küçük bir azalış olur. Stereosilya modiolusa doğru büküldüğünde direnç artar ve elektrik akımı azalır. Böylece EP'de artış olur. Bu şekildeki voltaj iniş-çıkışları, CM, dış hair cell'lerin varlığına bağlıdır. Nöral potansiyelden farklı olarak CM'nin dalga şekli basilletr membranın hareketini görüntüler.
Sumasyon potansiyeli (SP) kohleada seslere cevap oluşturan bir DC potansiyeli olarak kaydedilir. Bu DC potansiyelinin kayıtları scala timpani,media veya vestibulide yapılabilir ve hatta bazı koşullarda insanın kulak kanalındaki bir gross elektroddan yapılabilir. Buradaki potansiyel (+) veya (-) olabilir ve elektrod lokalizasyonu veya stimulus frekansı ve seviyesine bağlı olarak polariteyi ters çevirebilir. SP muhtemelen çeşitli orjinlere sahiptir. Fakat büyük ölçüde dış hair cell'lerin ve kısmende iç hair cell'lerin intrasellüler potansiyellerden çekilen stimuluslarm DC değişimlerini yansıtır.
Tüm sinir veya bileşik aksiyon potansiyeli (ÇAP) işitme sinir liflerinin tümünün deşarjından doğar. ÇAP yuvarlak pencere veya işitme siniri yakınında yerleşmiş bir gross elektrod ile ve yüksek frekanslı aniden ortaya çıkan sinyalleri kullanarak en etkin şekilde kaydedilir. Böyle sinyaller senkronize nöral aktiviteyi üretir. Bu aktivite de ÇAP dalga formu oluşturmak için toplanırlar. ÇAP klinik olarak skojp elektrodları veya dış meatustaki bir elektrod ile yada transtimpanik bir yaklaşımla yuvarlak pencere yakınına yerleştirilen elektrod ile kaydedilebilir. Son zamanlarda SP büyüklüğünün ÇAP büyüklüğüne oranı perilenfatik fistülanın bir göstergesi olarak kullanılmıştır. Fakat bu indikatörün geçerliliği hala şüphelidir.
İŞİTME SİNİRİ FİZYOLOJİSİ
İşitme siniri insanlarda yaklaşık 3000 .kedilerde 50000 kadar lif içerir. Son yıllarda en önemli araştırma sonuçlarından birisi; nöronların ( Tip l radial lifler ) % 90-95'inin iç hair cell'leri innerve etmesi, buna karşın %5-10 kadarının (Tip II dış spiral lifler) dış hair ceü'lerle ilişkili oluşunun gözlenmesidir. İşitme sinir liflerinden sağlanan kayıtların tümü değilse bile çoğu, iç hair celilerle ilişkili olan tip l liflerindendir.Bu radial lifler spiral ganglionda bipolar hücre yapılarına sahiptirler. Dış spiral lifler monopolardırlar ve myelinli değildirler.
İşitme sinir fonksiyonunun en temei ölçümleri spontan otenlar, ayar eğrileri ve yoğunluk (Oran-Düzey) fonksiyonlarıdır. Memelilerdeki işitme sinir liflerinin çoğu akustik stimülasyon olmadığında deşarj olurlar ve spontan deşarj oranlarına göre üç kategoriye ayrılmışlardır: Yüksek (18-120 spike/sn), orta (0.5-18 spike/sn), düşük (0-0.5 spike/sn).
Yüksek spontan aktivite oranları olan lifler diğer liflere göre daha düşük düzeylerdeki işitme sinyallerine cevap verirler. Diğer bir ifade ile en hassas lifler, en fazla spontan aktiviteye sahip olanlardır. Yüksek spontan oranlara sahip lifler, dış hair celilere dönük iç hair cell'ler tarafından sonlanırlar ve kalın dendritlere sahiptirler. Düşük ve orta spontan oranlara sahip olanlar ise ince dendritlere sahiptirler ve modiolusa bakan iç hair cell'ler tarafında son bulurlar.
Devam eden araştırmalar göstermiştir ki yüksek spontan aktivite değerlerine sahip lifler, diğer liflere göre kohlea nükleusunda farklı terminasyonlara sahiptirler.
Tek bir işitme sinir lifinin ayar eğrisi işitme siniri fonksiyonunun en temel göstergesidir. Ayar eğrishn keskin ucu lifin karakteristik frekansını (CF) tanımlar. Böylece düşük CF'si olan birimler kohleamn apikal bölgesindeki iç hair cell'leri innerve eden liflerdir. Yüksek CF'li lifler ise bazal bölgedeki iç hair cell'leri innerve ederler.
İŞİTME SANTRAL SİNİR SİSTEMİ
İşitme yolu canalis spiralis modioli içinde oturmuş olan ganglion spirale bipolar nöronlarının uzantılarından başlar.
Primer İşitme Lifleri: İşitme impulsları 8. sinirin pars kohlearis'i ile beyin sapına taşınır. İşitme impulslarının reseptörü kohlea içinde Corti organıdır. Corti organı içindeki hair cell'ler özel reseptör hücrelerdir. Bunlara ganglion spirale nöronlarının perierik uzantıları bağlanır. Ganglion spirale modiolus içinde bulunur. Bu ganglionun merkezi uzantıları N. cohlearisi yaparlar.
N.cohlearis lifleri ponsun altında, corpus restiforme'nin dış yüzünde bulunan, Nükleus cohlearis dorsalis ve Nükleus cohlearis ventraliste sonlanır. Bunlara primer işitme lifleri denir.
İşitme sisteminin tipik özelliklerinden biri bu sistemin her yerinde tonotopik bir lokalizsyonun bulunmasıdır. Kohlear nükleuslarda alçak tonlu sesler, ventral nükleus ve dorsal nükleusun ventral kısımlarına, yüksek tonlu sesler dorsal kohlear nükleusun arka kısımların giderler.
Sekonder İşitme Lifleri: kohlear nükleustan çıkan liflerdir. Bunlar 3 stria yaparlar.
a.) Stria acusticus ventralis: Ventral kohlear nükleustan çıkarak karşı yarıya geçerler. Pons tegmentumu'nun ön tarafında corpus trapezoidum'u oluştururlar. Karşı yarıda yükselirler. Bu yükselen liflere lemniscus lateralis denir. Liflerden bazıları, corpus trapezoideum nükleusları ve nükleus olivarius superior'da aynı ve karşı taraflı sonlanırlar.
b.) Stria acusticus dorsalis ve intermedius: Her ikisi de dorsal kohlear nükleustan ve ventral nükleusun arka kısmından çıkarlar Hepsi karşı tarafın lemniscus lateralisine giderler. Bütün sekonder lifler N. Olivarius superior, N.Tnapezoideum ve Formatio reticularis'te sonlanan lifler verirler.
Nükleus Olivarius Sup. ve N. corpus Trepezoideum'dan, çıkan liflere tersler işitme lifleri denir.Bunlar aynı taraflıdır. Böylece Lemniscus lateralis, sekonder ve tersier işitme liflerinden oluşur. Lemniscusun lifleri arasında bulunan N. lemniscus lateralis'lerde bazı lifler sonlanır veya bunlardan bazı yeni lifler lemniscus'a katılır. Lemniscus lateralis'ler, mezensefalon'da coliculus inferior'larda aynı taraflı veya karşı taraflı olarak sonlanırlar. Bazı lifler ise doğrudan brachium colliculi inferiordan, corpus geniculatum medialeye geçerler. Colliculul inferior'un kendi lifleride aynı yoldan corpus geniculatum medialeye geçerler, corpus geniculatum mediale lifleri ise tractus geniculo corticalis yoluyla, gyrus temporalis superior'daki 41 nolu işitme merkezine (Heschl) aksederler.
İşitme yolundaki aracı nöronların çokluğu nedeni ile reflex bağlantılar tam olarak anlaşılamamıştır. Ancak işitme korteksinden, kohleaya yükselen yol içinde geriye inen bir descedens yol vardır. Nükleus olivarius superior'dan çoğunluğu karşı , azınlığı aynı taraflı olmak üzere bazı lifler tekrar kohlaya geri dönerler. Bu lifler işitme siniri etkinliğini baskı altına alan bir refleks yolu yaparlar. Ayrıca aracı nöronlardan kohlear nükleuslara inen lifler işitme spektrumu içindeki bazı frekansları inhibe ederler. Buna işitmenin keskinleştirilmesi denir. (Örneğin : bir orkestrayı dinlerken topluce gelen enstrüman sesleri arasında belli bir enstrüman sesinin daha baskın olarak duyulması).
İşitme yolundan formatio retikülaris'e karışan lifler yüksek seste başın çevrilmesi ve gözlerin kapanmasını sağlar.
Colliculus inferior nöronları ses kaynağının lokalizasyonunu saptarlar. Bir taraf n.kohlearisin tahribi o tarafta tam sağırlığa yol açar
Lateral lemniscus lezyonlarm da iki taraflı işitme azalması ortaya çıkar. Bir taraf işitme korteksi çıkarılırsa karşı taraf ses lokalizasyonu bozulur.